Evolution of the Kidney-Juniper publishers
JUNIPER PUBLISHERS-OPEN ACCESS ANATOMY PHYSIOLOGY & BIOCHEMISTRY INTERNATIONAL JOURNAL

Aim of the work: This article will
elucidate the extraordinary parallels between the evolution of the
kidney from provertebrates to man. In invertebrates, the excretory
structures are classified into three types included contractile vacuoles
in protozoa, nephridia (flame cell system) in most invertebrate animals
and Malpighian tubules (arthropod kidney) in insects. While, in
vertebrates, there are three distinct excretory organs formed in
succession during the development of the vertebrate kidney, they are
called pronephros, mesonephros and metanephros.
Conclusion: From this review,
it can be concluded that the important factors in the evolution of the
basic structure and function of the vertebrate kidney appeared
associating with body fluid– regulation, involving the maintenance of a
constant water and salt content of the body. As the evolution of the
vertebrate kidney illustrates how pronephric, mesonephric and
metanephric kidneys are represented successful evolutionary responses to
the surrounding environmental pressures.
Keywords: Development of
kidney; Pronephros; Mesonephros; Metanephros; Opisthonephros;
Archinephors; Flame cells; Contractile vacuoles; Malpighian tubules
Introduction
Evolution of the kidney is a hot topic for many
researches and biologists as there is no better place to see the impact
of evolutionary pressures on organ development than in the kidney and to
study the ability of human metanephroi to differentiate after
transplantation into functional mature nephrons [1].
Aim of the work
All vertebrates have kidneys like the human kidneys,
they are made of many nephrons. However, there are many differences in
the structure and function of various vertebrate kidneys that adapt them
to the environment in which the animals live. This article will
elucidate the extraordinary parallels between the evolution of the
kidney from provertebrates to man.
The Excretory structures in Invertebrates and Vertebrates
The excretory system regulates the chemical
composition of body fluids by removing metabolic wastes and retaining
the proper amount of water, salts and nutrients. The invertebrate
excretory structures are classified in according to their marked
variations in the morphological structures into three types included
contractile vacuoles in protozoa, nephridia (flame cell system) in most
invertebrate animals and Malpighian tubules (arthropod kidney) in
insects [2].
There are three distinct excretory organs formed in
succession during the development of the vertebrate kidney, they are
called pronephros, mesonephros and metanephros. The pronephros is the most primitive one and exists as a functional kidney only in some of the lowest fishes and is called the archinephros. The mesonephros represents the functional excretory organs in anamniotes and called as opisthonephros. The metanephros
is the most caudally located of the excretory organs and the last to
appear, it represents the functional kidney in amniotes [2-4].
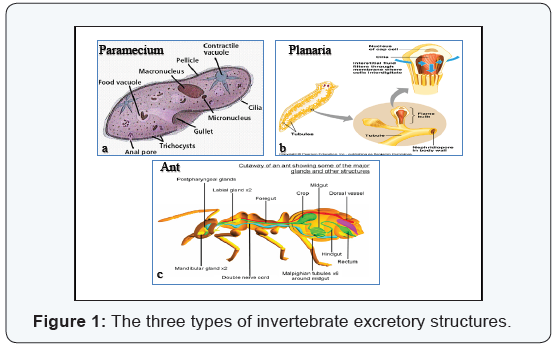
Invertebrate excretory structures
Contractile vacuole: The contractile
vacuole presents in the protozoa (Figure 1A), do not be considered as a
true excretory organ, as ammonia and nitrogenous wastes left the cell
by diffusion, while the contractile vacuole is a true organ for water
and salt balance. In amoeba proteus, the excess water is collected in
fine and many vesicles surrounding the membrane of the contractile
vacuole. These vesicles are emptied their contents into the vacuole.
This vacuole is moving inside the cytoplasm for a time until it reaches a
certain size where it is then passes up through the plasma membrane and
empties its contents into the surrounding medium through a small pore
in the plasma membrane. When the water is expelled from it, it starts to
refill immediately [2].
Nephridia (Flame cell system): In most of the invertebrates, excretory organs are called nephridia or nephridial tubules.
There are two types of nephridia, the first is protonephridium
and the second is metanephridium. The protonephridium had
a blind tubule system, it is present in many invertebrates such as
flatworms. At the end of each blind tubule of the nephridium is a
ciliated flame cell (Figure 1B). The fluid enters the system through
the flame cells and then passes down through the tubule where
some ions and molecules are reabsorbed. The wastes are expelled
outside through excretory pores on the body surface [2-4]. The
metanephridium or called true nephridium has an open tubule
system which is surrounding by a vascular network. It is present
in some invertebrate such as the earthworm. In the earthworm,
each nephridial tubule occupies two adjacent segments. The
tubule is connected to the coelom at one end by a ciliated funnel,
or nephrostome, and at the other end is opened to the exterior by
an excretory pore called nephridipore.
In amphioxus (primitive chordates), there is a series of
excretory tubules opened into the atrium or pericardial space.
Each lies on the outer dorsal side of the secondary gill bar. They
were apparently of ectodermal origin, has no connection with
the coelom and are composed of numerous flame cells called
“solenocytes” which collected wastes. The solenocytes are
attached to the walls of blood vessels and are bathed by coelomic
fluid. Those belonging to a given tubule entered a common
excretory canal, which in turn is opened into the atrium through a
small excretory pore, is called a nephridipore [2,3].
Malpighian tubules (Arthropod kidney): The insects have
a special excretory system which is formed of Malpighian tubules
and rectal glands (Figure 1C). The Malpighian tubules are thin,
delicate, convoluted and blind. They have no blood supply. The
body fluids are drawn into the tubules by osmosis due to the
large concentrations of potassium inside these tubules, and then
the body fluids are passed back into the body and reabsorbed by
the rectal glands. While the nitrogenous wastes emptied into the
insect’s gut. This excretory system conserved water and is suitable
for insects which lived in a dry environment [2] (Figure 1).
Evolution of the vertebrate kidney
Evolution of the vertebrates is a fascinating story viewed
in terms of the external osmotic environment in which various classes evolved. Fresh water, marine and terrestrial habitats
possessed different problems for the maintenance of internal
water balance and the excretion of nitrogenous wastes. The
evolution of the kidney in vertebrates illustrates how pronephric,
mesonephric and metanephric kidney, represent successful
evolutionary responses to these environmental pressures. So
many variations in the evolution of the kidney are correlated
with these environmental factors. Variations in the structure of
the vertebrate kidney from fish to man are primarily in the nature
of alterations in number, complexity, arrangement and location of
the kidney tubules [5,6].
Embryological origin: The kidney in all vertebrate is
originated from the intermediate mesoderm. The mesoderm
which will form the kidney was called nephrogenic mesoderm.
The kidney as a whole is made up of two elements, the kidney
duct and the kidney tubules. The kidney tubules called nephrons,
they are evolutionary modifications of the nephridia and they
are the kidney’s functional units. The development of the kidney
is somewhat complex in that two or three different kidneys
(depending on species) formed in temporal and spatial sequence.
The first, most anterior and largest to develop is the pronenphric
kidney. The second kidney to form is the mesonephric kidney. In
birds, reptiles and mammals, a third kidney developed posterior
to the mesonephros, called the metanephric kidney [3-6].
Effect of environment on the nephron structure and
function: The components of the nephron in higher vertebrates
and man are the glomerulus, the Bowman’s capsule, the proximal
convoluted tubule, the loop of Henle and the distal convoluted
tubule. The glomerulus filters the blood. The Bowman’s capsule
also filters the blood and their cells (podocytes) prevent the
passage of the large molecules as blood proteins and blood cells to
the Bowman’s space. The proximal convoluted tubule reabsorbs
water, salts, glucose and amino acids. Loop of Henle reabsorbs
water and small molecules. The distal convoluted tubule secretes
H2 ions, potassium and certain drugs [5,6].
However, there are three types of nephrons in different
species of vertebrates. The first type is presumably the most
primitive, found in amphibians, fresh water, bony fishes and
elasmobranches, there is a renal corpuscle of good size and
hence a high water output. The second type was that found in
many marine teleosts and in reptiles, the corpuscle is small or
absent with shortening of the renal tubule, hence increases salt
excretion and water conservation (water output is low). The third
type is seen in mammals and in a less extreme form, in birds.
The glomerulus is large, but there is interjected into the middle
of the convoluted tubule, a long slim extra- segment, the loop of
Henle. This slim segment appeared to be a powerful resorption
of water, hence despite a plentiful output at glomerulus, relatively
little water reached the bladder [5]. The distribution of the
three types of nephrons among vertebrates is correlated with
the environmental factors. These variations can be explained as
followed:
- The first type is present in early fresh water vertebrates. Such an animal lived in a medium more dilute than its own fluids and hence is in danger for overdilution of these fluids by osmosis through the surface of the body and of the gut. To prevent this, large amount of water should be eliminated and this is afforded by the presence of a large corpuscle.
- The second type is present in marine teleosts where high salinity of the surrounding medium is present. In this type, water should be conserved and much salt eliminated. So the glomeruli are frequently reduced or absent. The aglomerular kidney is present in seahorses and pipefishes [7]. So water output consequently reduced, further salts and wastes are excreted by the cells of the gills membranes and rectal glands as well as by the kidneys. The terrestrial vertebrates, lived in a dry environment have the same problems as a marine fish, water should be conserved. In modern reptiles, this is accomplished by reduction in size of the renal corpuscles with a consequent decrease in water output.
- The third type is present in birds and mammals. These animals developed a different method of conservation of water. There is a normal glomerulus of large size and consequent high water output. The complex tubule and the presence of loop of Henle result in absorption of much of the water, the product is a relatively concentrated urine [5,6].
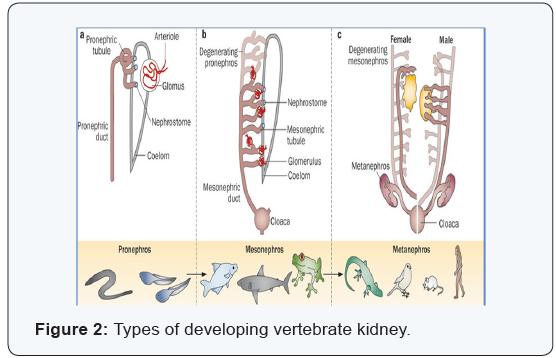
Archinephors: The primitive
vertebrates possess an
excretory organ which is referred to as an archinephros or
holonephros. This is consisted of a pair of archinephric ducts
located on the dorsal side of the body cavity and is extending the
length of the coelom. Each duct is joined by a series of segmentally
arranged tubules, one pair of tubules to a segment. At its other
end, the tubule is opened into the coelom by a ciliated, funnelshaped,
peritoneal opening called the nephrostome. Also another
structure formed in connection with each tubule, it was also
called external glomerulus, a small knot or cluster of capillaries
interposed within the course of an arteriole and located in
close proximity to the nephrostome. Tissue fluids, exuded at the
glomeruli, are passed in the coelom and hence the nephrostomes into the
kidney tubules and finally through the archinephric ducts
to the outside [8].
The Anamniote kidney: The developing kidney in an amniotes
differentiated in two stages: pronephros and opisthonephros.
Pronephros: The pronephros is actually consisted of a varying
number of anteriorly located pronephric tubules together with
a pair of archinephric ducts duct (which called here pronephric
duct). The tubules and ducts lay in the dorsolateral mesoderm on
either side of the mesentry that supported the gut. The tubules
were segmentally arranged, connected with the near pronephric
duct at its anterior end. The outer end of the tubules opens into
the coelom by means of nephrostomes. The nephrostome and the
part of the tubule near it are ciliated. Most forms are possessed
internal glomeruli. These are knots of interarterial capillaries,
each surrounded by a double wall structure called Bowman’s
capsule, the two together are known as renal or Malpighian
corpuscle. Sometimes, several glomeruli united to form a large
glomus. In some cases, pronephric tubules expanded so as to
form pronephric chambers or one large pronephric chamber.
The pronephros was degenerated soon after it have been formed
[2,3,5,6].
Opisthonephros: Since the pronephros in most cases
is a transient structure, the opisthonephros is served as the
adult kidney in lampreys, most fishes and amphibians. In many
accounts, the term mesonephros is used in describing what they
are here calling the opisthonephros. Biologists have realized that
the opisthonephros of cyclostomes, fishes and amphibians is not
quite comparable to the mesonephros of embryonic amniotes
even though the two are structurally similar in many ways.
It is important to mention that the opisthonephros of
anamniotes actually extended over a region which in amniotes
would form the mesonephros and metanephros . In forms
possessed an opisthonephros, there is a general tendency toward
a concentration of kidney tubules toward the posterior end of
the organ. The anterior portion frequently losses its significance
as an excretory organs and in the male, may became part of the
reproductive system. Furthermore, the connection of the kidney
tubules with the coelom is lost in most case and the presence of
renal corpuscles with internal glomeruli being typical [6,8].
Structure of the opisthonephros
Each tubule differentiated into a narrow neck at the end of
the renal corpuscle, followed in turn by secretory and collecting
portions. The collecting portion connected with the archinephric
duct. The secretory part of the tubule forms two loops named the
proximal and distal convoluted segments or tubules [3,9].
The Amniote kidney
In reptiles, birds and mammals including humans, three
types of kidneys are recognized, pronephros, mesonephros
and metanephros. These are appeared in succession during
embryonic development, but only one, the mentanephros is persisted to become the functional adult kidney. Mesonephros
and metanephros are actually represented different levels of the
opisthonephros of the anamniotes, the metanephros being the
equivalent of the posterior portion. In all forms, an anteriorly
located pronephros is present during early stages of development,
but it is soon degenerated and the more posterior mesonephros
then is developed. The duct of the pronephros is persisted to
become the duct of mesonephros. This is actually the same as the
archinephric duct. The mesonephros is persisted for a time and
then is degenerated. In the meantime, the metanephros developed
from the region posterior to the mesonephros. Portions of the
mesonephros persisted to contribute to the reproductive system
in the male or to remain as mere vestigial structures, without any
apparent function in female [10].
Pronephros: The pronephros in amniotes is formed in a
manner similar to that of anamniotes. Segmentally arranged
pronephric tubules appeared in the intermediate cell mass in
some of the anterior segments of the body. They are first solid
structures, but they soon are hollowed out, one end established
a connection with the coelom. The tubules are appeared in
succession in a cranio-caudal direction. A variable number of such
tubules are formed in different species. In the chick, for example,
10 or 11 pronephric tubules are formed on each side from the 5th
to the 15th or 16th segment, the last tubule is enlarged caudally and
established a connection with the cloaca. Thus, a long pronephric
duct is formed, the anterior end of which is connected to a series
of tubules with coelomic connections. The tubules soon are
disappeared, in fact, the anterior tubules may degenerate before
the posterior ones even formed. External glomeruli may or may
not form. So, in mammals, pronephric tubules are appeared
only as the merest of vestiges. Hence, the pronephric duct can
scarcely be said to be formed as the result of fusion of tubules.
Nevertheless, it is appeared in the nephrotome region, first as a
solid cord which grow back to the cloaca , hollowing out to become
a typical pronephric duct [4,10].
Mesonephros: The mesonephros of amniote embryos
have essentially the same structure as the kidneys of fishes and
amphibians except that nephrostomes are rudimentary in most
birds and seldom appeared in mammals. In embryonic chick,
the mesonephros reaches its peak of development at the 11th day
of incubation, halfway through embryonic life. In mammals, it
reaches its peak earlier at the 9th week of gestation. In a human
fetus, it is appeared after 4 weeks of embryonic life (20- somites
stage). A wave of differentiation is occurred along the nephrogenic
mesoderm, so that even before the last mesonephric tubules at
the caudal end of the series has been formed, the earliest ones
at the anterior end has been already involuted. The result is
that at peak development of the human mesonephros, there
are about 30 functioning renal corpuscles, although as many as
80 have been formed by that time. The mesonephroi of various
species of mammals differ in the number of mesonephric tubules
formed. Those in man, cats and guinea pigs are relatively small as compared with the mesonephroi of rabbits [4,10].
Fate of Mesonephros: Although the mesonephros is basically
an embryonic kidney in amniotes, it functions for a short time
after birth in reptiles, monotremes and marsupials. In the
meantime, a new kidney to be used by the amniotes the rest of
life, the metanephros, is in the process of development. When the
metanephros took over the functions of a kidney, the mesonephros
involuted and only remenants remained after birth [10].
Mesonephric remnants in adult amniotes: Small remnants
of the mesonephroi are remained in both sexes after the
metanephroi involuted. In mammals, the remnants consisted
of groups of blind tubules known as the paradidymis and the
appendix of the epididymis located near the epididymis and as the
epoophoron and paroophoron near the ovary. The mesonephric
ducts remained as sperm ducts in male amniotes, but they are
involuted in females and thus remained only as short, blind
Gartner’s ducts coursing in the mesentry of the oviducts [10,11].
Function of mesonephric tubules: The function of
mesonephric tubules much like the nephrons of the adult kidney.
A filtrate of blood from the glomerulus enters the capsule and
flow into the tubule, where selective resorption of ions and
other substances are occurred. A major difference between
the mesonephric kidney and the permanent kidney of higher
vertebrates is the relative inability of the mesonephros to
concentrate urine. This is related to the elongated structure of the
mesonephros and the absence of a well developed renal medulla,
a structural adaptation of land animals to preserve water by
concentrating it through an elaborate countercurrent exchange
mechanism.
Such a fluid conserving mechanism was not needed by the
embryo which lived in a bath of amniotic fluid, just as preservation
of body water was not a problem for the mesonephric kidney of
fishes and aquatic amphibians [10].
Metanephros
The metanephros is found only in amniotes and human, is
arising posterior to the mesonephros on each side and is more
compact than the latter organ. It comes from a level which is
corresponded to the most posterior portion of the opisthonephros
of the anamniotes. The development of the metanephros begins
with the appearance of a tiny bud-like outgrowth from the
mesonephric duct just cephalic to the point where the duct
opened into the cloaca. The outgrowth, is called the metanephric
diverticulum. This diverticulum is pushed into the posterior
portion of the intermediate mesoderm, which is condensed around
the diverticulum like a cap to form the metanephric blastema.
Thus, the metanephric kidney had a dual origin, the metanephric
diverticulum, which gives rise to the ureter, the renal pelvis and
the collecting duct system and the intermediate mesoderm from
which the tubular units of the kidney arose [11-13].
Differentiation of the metanephric tubules
The metanephric blastema organize the metanephric tubules,
they commence as S-shaped tubules. The upper arm of each
tubule grows toward and finally opens into a collecting tubule.
The lower arm is invaginated by a developing glomerulus to
become a Bowman’s capsule. The mammalian metanephros
exhibits a greater organization than that of lower amniotes.
The organization is the result of the formation of a long thin,
U- shaped loop of Henle interposed between the proximal and
distal convoluted tubules. As the loops of Henle elongated, they
grow away from the surface of the kidney and toward the renal
pelvis. The kidney therefore consisted of a cortex in which are
concentrated the renal corpuscles and a medulla which consists
of the hundreds of thousands of loops of Henle and common
collecting tubules.
The loops of Henle and collecting tubules give the medulla
a striated appearance in frontal section. They are aggregated
into one or several conical lobes (pyramids), depending on the
species. The pyramids are tapered to a bunt apex (renal papilla),
projecting into the renal pelvis and are surrounded by extensions
(the calyces) of the pelvis. Each collecting tubule drains a small
number of metanephric tubules and then empties into the renal
pelvis near the end of the papilla.
The metanephric tubules of reptiles have no loop of Henle,
and those of birds have only a very short equivalent segment. So
reabsorption of water in reptiles and birds also are occurred in
the cloaca, into which the ureters are opened. The glomeruli in
reptiles and birds are reduced in size and exhibited only two or
three short vascular looped within Bowman’s capsules [11-13].
Comparative anatomy of metanephros: The comparative
anatomy of metanephros in amniotes are described [3,6,14].
In reptiles, the kidneys are restricted to the posterior half
of the abdominal cavity and are usually confined to the pelvic
region. They are generally small and compact but their surfaces
are lobulated. They are elongated or slender in shape.
In birds, the kidneys were situated in the pelvic region of the
body cavity and the two frequently united at their posterior ends.
They are flat–shaped. Their surface is lobulated, deep fissures
between the lobules are present, serving for the passage of the
branches of the renal veins.
In mammals and human, the typical kidneys are compact beanshaped
attached to the dorsal body wall. They are retroperitoneal.
The ureter leaves the medial side at a depression called the hilum.
At this point, a renal vein also leaves the kidney and a renal artery
and nerves enter it. The metanephros is surrounded by a capsule
of connective tissue under which lie the cortex.
Lobulation of vertebrate kidneys
Many metanephric kidneys are lobulated as in reptiles, birds
and others,each lobe consisted of clusters of many tubules.
Lobulation is occurred also in human infants but later is
disappeared during the first year after birth. Varying degrees of
lobulation, however, on occasion, persisted through life [4].
Arterial supply of the vertebrate kidneys
In reptiles and birds, the arterial blood supply to the
metanephric kidney, is via a series of two or more renal arteries
of segmental origin. In mammals, there is usually a single renal
artery, but it very often bifurcated before reaching the kidney.
Upon entering the kidney of mammals, the renal artery divides
into humerous branches which passed radially toward the cortex
as interlobar arteries. At the base of the cortex, the interlobular
arteries give off arcuate arteries which arch along the base of
the cortex more or less parallel to the surface of the kidney.
From the arcuate arteries arise tiny interlobular arteries which
in turn, give off afferent glomerular arterioles terminating in
glomeruli. Emerging from each glomerulus is an efferent arteriole
which passes directly to a capillary bed surrounding the tubule.
Emerging from the capillaries are arcuate veins. The kidney is
drained by one or several renal veins.
The kidney of reptiles and monotremes is similar to a slight
degree, to that of birds, also has an afferent venous supply via
a renal portal system. The portal vessels terminate the same
peritubular capillary beds as do the efferent glomerular arterioles
[3,11].
Conclusion
From this review, it can be concluded that the important
factors in the evolution of the basic structure and function of the
vertebrate kidney appeared to have been associated with body
fluid – regulation, involving the maintenance of a constant water
and salt content of the body. As the evolution of the vertebrate
kidney illustrates how pronephric, mesonephric and metanephric
kidneys are represented successful evolutionary responses to the
surrounding environmental pressures.
For
more Open Access Journals in Juniper Publishers please
click on: https://juniperpublishers.com
For more articles in Anatomy Physiology & Biochemistry International Journal please click
on: https://juniperpublishers.com/apbij/index.php
For more Open Access Journals please click on: https://juniperpublishers.com
To know more about Juniper Publishers please click on: https://juniperpublishers.business.site/
For more articles in Anatomy Physiology & Biochemistry International Journal please click
on: https://juniperpublishers.com/apbij/index.php
For more Open Access Journals please click on: https://juniperpublishers.com
To know more about Juniper Publishers please click on: https://juniperpublishers.business.site/

Comments
Post a Comment